从万有引力到经济引力(1)
——试论财富的分配规律
张国文
(武汉市洪山区人民政府
武汉 430070)E-mail:
mengke130@sohu.com
2004.08公布于潜科学网站
内容摘要:本文用经济学和生物学的观点考察无机界,用物理学的方法研究生物界和人类社会,通过对万有引力定律等物理定律的综合研究、拓展,导出了生物界和人类社会的引力定律,揭示了社会财富的分配规律,同时也为人们探索和了解无机界、生物界和人类社会之间的本质联系提供了一种新的理论方法和途径。
关键词:
力学 万有引力 聚财力 经济运动1
、前言人类及其社会是自然界不可分割的一部分,物质运动是生物及其经济运动的基础,生物和经济运动是物质运动在更高层次的表现形式。19世纪,马克思就曾在他的《资本论》序言中认为,经济社会形态的发展是一个自然历史过程。在另一篇文章中,马克思写道:“科学,只有从自然科学出发,才是现实的科学。历史本身是自然史的,即自然界成为人这一过程的现实部分。”[1] 因此,历代经济学家都喜欢借用自然科学的成果来解决经济学的问题。如亚当.斯密的《哲学探讨的指导原理:以天文学史为例》,著名经济学家乔治斯库.罗根的《熵定律与经济问题》(1971)都是基于物理理论的经济论文。[2]
力是自然科学理论大厦的基石;力学理论的建立是自然科学从幼稚走向成熟的重要标志。因此,经济学家尤其垂青于自然科学中的力学。早在19世纪初叶,空想社会主义者傅立叶和欧文等人就曾明确指出,在人类社会中,有着类似于自然界的万有引力的东西在支配着社会的运动。傅立叶将这一支配社会运动的引力称为情欲引力。他认为,情欲引力就是万有引力在社会经济运动中的表现形式。他说:“情欲引力的规律在各个方面都符合牛顿和莱布尼茨所阐明的物质引力规律。物质世界和精神世界在运动体系上具有统一性。”[3]经济学家帕累托则认为:“理论经济学就是力学或类似于力学。”[4]
虽然许多经济学家都感觉到社会经济运动中有一种类似万有引力的力存在,但是,至今也没有那个经济学家能够真正找到这种力。当然,在经济学中,也有一些力的概念,例如生产力、劳动力、竞争力等等。但它们并未像物理学中的力那样,成为整个学科的基石。原因在于,所有这些力都是抽象和模糊的。它们既没有从定性上给人们提供定义清楚的概念,也不能从定量上做出什么描述。因此,要让力学成为经济学的基础,或者说要使经济学与物理学相融通,除非在经济学中定义或找出新的力。
本文从研究万有引力入手,将万有引力定律和库仑定律进行综合后运用到人类社会的经济运动,导出了社会经济运动中的引力公式,由此提出了一种新的财富分配理论。
2、自然力
2.1、万有引力
牛顿(Isaac Newton)(1686)发现的万有引力定律的一般表述是:任何两个物体之间都有相互吸引力,这个吸引力的大小与两物体的质量成正比,而与它们之间的距离的平方成反比。
如果用m1、m2表示两物体的质量,用r表示它们之间的距离,则两物体之间的吸引力F的大小为:
F=Gm1m2/r2 G为引力常数。
万有引力定律是一个天才的发现,但是三百多年来,人们对万有引力定律的认识、理解和应用都存在着较大的片面性,以至于在一个相当长的时间内,许多人误以为,我们的宇宙就像一座上足了发条而精确运转的机械钟,知道了它的现在,就可以毫不费力地推导出它的过去,预测到它的未来。其实,这是对牛顿万有引力定律的误解。我们知道,万有引力定律揭示了一切物质之间都存在着吸引力这一自然界的内在本质。这种力一方面为天体的运动提供了向心力源,使月球围绕地球转,地球围绕太阳转,太阳绕着银河中心转,让宇宙始终保持和谐与秩序,另一方面它在恒星、行星和卫星形成之时,帮助它们积聚了物质,为宇宙间的竞争提供了动力。
今天,各种恒星、行星和卫星及其它一切天体仍然在利用万有引力吞食广袤太空中的各种质量——陨星、宇宙尘埃等宇宙漂泊者。宇宙中的各种天体,除了在按照笛卡尔和拉普拉斯的描述作机械运动外,还在进行着有物质和能量交换的非线性运动。同时,从根本上讲,宇宙是复杂的多体系统,不存在单纯的二体系统。牛顿万有引力定律的二体表述式只是对天体运动的一种近似描述。因此,要使牛顿万有引力定律具有普遍的意义,就必须将其改造成能描述多体系统的一般形式。在这里,我们先不考虑宇宙的动态运动,用静力学来分析一个多体系统的受力情况。
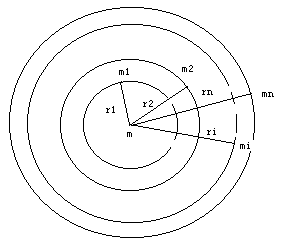
设在某一质量为m的物体周围,有n个物体,它们的质量分别为m1、m2、…mi…mn,它们与物体m的距离分别为r1、r2、…ri…rn(如图),那么根据牛顿万有引力定律,物体
图:质量为m的物体周围的n个物体在平面上的投影
m所受的总的吸引力的大小就是:
F=Gm1m/r12+Gm2m/r22+…+Gmim/ri2+…+Gmnm/rn2
或者
F=Gm∑(mi/ri2) …………………………①
这里,∑(mi/ri2)=m1/r12+m2/r22+…+mi/ri2+…+mn/rn2
可将①式改写成F=4πGm∑(mi/4πri2)(等式右边的分子和分母同时乘以4π)。或 F=ξm∑(mi/4πri2) ……………………②
这里ξ=4πG,4πri2是半径为ri的球面面积,mi/4πri2 表示单位面积所含的质量数。因此,从静力学的角度看,m物体所受的吸引力的大小,实际上与周围环境的质量分布状况有关,单位面积质量分布越多,物体所受的吸引力就越大。所以,一个物体的受力状况与其周围的物体的分布状况有密切的关系。周围的物体越多,质量越大,该物体受力就越大。这里,我们所研究的物体的受力,是一种感受到的力,而不是物体所受力的失量之和的表现,即物体所受的总的合力的大小,等于所有力的绝对值的和,而不是各个力的矢量相加。
另外,由于周围物体对m物体的吸引力与m物体对周围物体的吸引力大小是相等的,所以,上述①式也反映了m物体对周围各物体的吸引力。这一点非常重要,因为它包含着深刻的经济学意义。在后面的讨论中,尤其是在讨论经济问题时,我们一般说被研究对象对环境各物的吸引力,而不说环境各物对研究对象的吸引力。
至此,我们对万有引力定律进行了改造,并做出了新的解释。它反映了宇宙的另一种运动——物质的聚集、分配和交换运动。现代天文学告诉我们,宇宙没有一刻是平静的,各种天体都在不断地与外界交换物质或者能量。几乎每时每刻都有星系或者超新星在爆炸瓦解以及彗星和小行星被大的天体吞食;同时,也不断地有新的恒星、行星、卫星和彗星在形成。总之,如果我们用经济学的观点来看宇宙天体的运动,就会发现天体对质量的占有(吸引)欲望比之人类对金钱或者财富的占有欲望,简直有过之而无不及。
2.2、库仑力
库仑(Charles-Augustin Coulomb)定律现在的一般表述是:在真空中,两个电荷之间的相互作用力的大小,与它们所带电量的乘积成正比,与它们的距离的平方成反比。如果用q1、q2分别表示两个点电荷所带的电量,用r表示它们之间的距离,那么库仑定律就可以用下列式子表示:
F=Kq1q2/r2这里K为比例常数。
如果在两个点电荷之间加入介质,那么上述库仑定律就变成下述表达形式:
F=q1q2/4πεr2(这里ε为介电常数)。
假如在电荷q周围有n个电荷,它们所带的电量分别为q1、q2、…qi…qn,它们与电荷q之间的距离分别为r1、r2、…ri…rn,那么根据前面对万有引力定律的改造办法,同理可以得到:
F=q∑(qi/εi4πri2) …………………………③
其中εi是电荷qi所在介质的介电常数。
从万有引力定律和库仑定律可以看出,它们有着完全相同的表达形式。这暗示了电磁力与万有引力之间存在着内在的一致性,为人们进一步地揭示自然力之间的内在联系提供了重要线索。事实上,库仑在发现该定律时,正是受到了牛顿万有引力定律的启发。
对于并不相互接触的两个物体或电荷,它们之间能产生相互作用力,当初人们很难理解。后来,法拉第把场和力线的观念引入物理学,使人们对力的媒递作用有了新的认识。法拉第认为,电磁力从电荷或磁极出发的传播,类似水面波纹的振动或空气粒子的声振动。当时,人们还不知道电磁波的存在,法拉第具有如此深邃的洞察力实属难得。现在人们都确信电荷对电荷的作用,是通过电场来传递的,或者说是通过电磁波这种媒介来传递的。不论是场还是电磁波,从本质上讲都是一种信息。自然,电荷之间传递的电磁波越多,相互作用力就越大。假如我们在两个电荷之间加入某一种介质,那么介质就会妨碍电荷相互交换电磁波,使电荷之间的作用力减小。因此,介电常数实际反映的是电荷电场信息被阻隔的程度。介电常数愈小,说明信息被阻隔的程度愈弱,电荷同周围电荷的作用力就愈强;介电常数越大,说明信息被阻隔的程度越强,电荷同周围电荷的作用力就越弱。例如,在真空中,电场信息被阻隔的程度最小,电荷对电荷的吸引力或者斥力最强;如果在两个电荷之间加入一种介质(如空气、塑料等),则电荷的电场信息会被阻隔一部分,电荷之间的作用力就相应地减弱;如果在两个电荷之间加入一介电常数为无穷大的绝缘体,则电场信息会被完全隔断,两个电荷之间的作用力(吸引力或者斥力)就为零。
由此可见,电场或者电磁波其实就是一种信息。电场的强弱实际上就是信息量的多少的反映。介电常数对电场信息的阻隔,如同隔音板对声音的阻隔一样。
3、生态系统中的引力
3.1、捕食力
动物的身体是由物质分子组成的,所以毫无疑问,动物与其他一切物体一样也存在着万有引力。那么除了这种可称量的体重(万有引力)外,动物还有没有另一种形式的吸引力呢?或者说在动物界有没有一种左右动物行为的力存在呢?
只要留心观察一下周围,我们就会相信动物界确实有这样的力存在。飞蛾扑火就是一个例子。这里火就是一个吸引源,而那些飞蛾则像一些没有生命的“粒子”,情不自禁地沿着“光场”运动到吸引源。
事实上,许多昆虫都有类似飞蛾的运动倾向。倘若我们把黑夜里的灯光和信息素看成是一种类似于电场或引力场之类的作用场,那么这些昆虫的运动(飞行)就与电子等带电粒子在电场或引力场中的运动没有多大的差别。如果用一种半透明的薄膜将灯光罩住,光线就会减弱,辐射的范围也将会缩小,这时“扑火的飞蛾”也会相应地减少;如果用一块黑色的厚布将灯光罩住,光线就不能透射出来,飞蛾见不到任何光线,也不会飞来扑火。这里光也是一种电磁波,薄膜和黑布也是一种电介质。它们对光线的阻隔,就是对光源信息的阻隔。
除了光可做动物的吸引源外,还有食物、性等也可作为动物吸引源。
在生态系统中各种生物或物体都是相互联系的,它们之间都充斥着相互作用,都存在着力。前面我们已经讲过,作为自然力,每一种力都应该有其作用的根源。
既然食物和性对动物有着强烈的吸引力,那么我们能否仿照上述四种作用,把食物(或性)等也作为一种“荷”,在动物界建立某种力的作用关系呢?
为了考察和研究动物生存、运动与食物的关系,同时考虑前面所讲到的引力对动物的作用,这里我们首先引进一个概念——捕食力,在本文中,捕食力是指动物在某一时刻捕获到食物的能力。捕食力实际上是一种引力。这种力的作用效应与万有引力和电磁力既有相同的地方,又有不一样的方面。捕食力是单向的或者说是反向的,只有被食者对捕食者有吸引力,而捕食者对被食者则表现为斥力。当然,这种效应并不防碍这种作用作为力的一种形式而存在。因为弱相互作用力也不是单纯的作用效应,它甚至没有通常的力所表现出的吸引和排斥作用。
捕食力的“荷”是食物,捕食力的作用“媒介”粒子是光子(可见光)和分子(传递气味的化学分子和传递声波的空气分子),其作用范围就是一般动物能够看见或听见或用嗅觉感觉到的距离。
对动物的行为进行深入研究就会发现,在一个生态系统中,某种动物(或动物社群等)的捕食力与下列一些因素有关:
(1)动物的特性、体质和捕食欲望。
动物的特性是指动物自身所独有的个性。它与动物在食物链中所处的位置密切相关。不同种类的动物其特性不同,捕食的对象和数量也相差悬殊。兔子以草为食,燕子以昆虫为食,老虎以肉(动物的尸体)为食。由于老虎处在食物链的最顶端,而牛处在食物链的中游,所以,即使牛的体重比老虎重,个头比老虎大,牛也只能被老虎捕食,而不敢去捕食老虎。因此,在一个食物链中,越是处在“营养金字塔”上部的动物,其捕食力就越强。
由此可以看出,动物的特性对动物的捕食力的影响有着决定性的作用。
动物的体质和捕食欲望对捕食力的影响,也是一个不可忽视的重要因素。体质好、食欲旺的动物,其捕食能力也强。反之,体质差、食欲弱的动物,其捕食能力则较弱。处在青壮年时期的动物体强身壮,食欲也旺,它们会捕食更多的食物;而一些老弱病残的动物,其体力不支,食欲不旺,既吃得不多,又难以捕到食物。
如果用F来表示动物的捕食力,用k来表示动物的特性、体质和捕食欲望,并约定动物在“营养金字塔”中的位置越高、动物体质越好、捕食欲望越强,k值就越大,那么很显然,动物的捕食力F与k成正比。即F∝k
值得说明一下的是,根据我们上述的规定,即动物在“营养金字塔”中的位置越高,k值就越大,显然有:
老虎的k值>狐狸的k值>鹿的k值>兔子的k值;
或者:肉食动物的k值>植食动物的k值>植物的k值。
(2)动物的体重。
生物学家研究发现,动物的进食量基本上与其体重成正比。在一个生态系统中,一个群落的捕食量也基本上与整个群落的体重成正比。这是因为动物的基本代谢是随动物的体重的增加而增加的。Ruhestoffwechsel研究认为,动物的基础代谢与其体重存在下述关系:[5]
单位时间的基础代谢(V)=N·Wb
这里N是比例常数,W是动物的体重,b为表面积的大小,其值接近1。除了极少数动物在及其特殊的情况下(如在冬季到来之前)会储藏一些食物外,动物一般都不储藏食物,它们需要多少食物就捕食多少食物,或者说它们捕食多少食物就吃多少食物。而动物的代谢量或者进食量是靠捕食力来保证的。基于这一事实,我们也可以说动物的捕食力与其体重成正比。这一点对于同种动物或习性相近的动物来说是很好理解的,我们常说:“大鱼吃小鱼,小鱼吃虾米,虾米吃淤泥。”这里起决定作用的是体重。
因此,如果我们用Q来表示动物的体重,那么在动物其他的条件完全相同的情况下,就有:F∝Q
(3)周围环境中食物的分布状况。
生物学家发现,在一个生态系统中,当单位面积的食物数量增加时,捕食者会作出两种反应:[6]
首先是数值上的反应,即单位面积里捕食者的数量会随食物的增加而增加。在自然界中,捕食者的数量是通过迁入或繁殖而增加的。当某一地区食物的数量增加时,一方面会吸引邻近食物匮乏地区的动物蜂拥而至,另一方面则会加速该地区动物的繁殖。前者是该地区原有群落之外的捕食力,它早己存在于生态系统之中,后者则是该地区原有群落中自生的新的捕食力。
在自然界中,只要食物有保证,生物就会迅速繁殖。而当食物的供应严重不足时,动物的繁殖就会下降或停止,从而导致群落个体的数量减少。我国学者裴新澍认为,以植物为食的鸟类、昆虫和其它动物,除了恶劣气候条件大量伤亡外,对于它们群体大小起调节作用的主要是食物的多少。赖克(Lack)(1954)在对鸟类的研究中发现,一般成鸟很少死于疾病或被捕杀,其数量的减少主要是受食物的影响,而在食物充足的地区鸟的数量较多。[7]
其次,是功能反应,即当单位面积食物数量增加时,捕食者会比在正常情况下捕食更多的食物。这种效应在许多生态系统中都十分明显。在自然条件下,由于动物都具有极强的繁殖能力,而环境能够提供的食物则是有限的,所以,在一般的情况下,虽然绝大多数动物都在不断地寻找和捕食食物,但它们还是经常处在饥饿或半饥饿状态。因此,当食物增加时,它们就会加紧进食以饱肚福,并加快繁殖后代。
由此可见,周围环境的食物数量增加,动物的捕食力也会增强。换言之,就是动物的捕食力与周围环境的食物数量成正比。
如果用Q来表示在捕食者周围r米远处的一个球面上的食物数量,那么在距离捕食者r米的球面上食物的分布面密度则为:Q/4πr2。
一般说来,环境是立体的(球体状的),而不是球面状的。所以食物的分布应该是在整个球体内,而不是只在一个球面上,但是我们可以利用微分法把球体分成无数个球面,这无数个球面到捕食者的距离分别为r1、r2、…ri…,各个球面上的食物分别为Q1,Q2,…Qi…,则这些球面上食物的面密度分别为:
Q1/4πr12,Q2/4πr22,…Qi/4πri2…。
因此,整个环境的食物的分布状况可由下式来表示:
∑(Qi/4πri2)=Q1/4πr12+Q2/4πr22+…+Qi/4πri2+…。
根据上面的讨论,显然有F∝∑(Qi/4πri2)。
(4)环境阻力和有关食物的信息。
所谓环境,就是动物所栖息的空间及其变化的各种条件或因素。如山川、草地、森林、海洋、植物、动物、气候、阳光等等。环境是动物赖以生存的基础,同时它对动物的生存也有制约作用。
气候条件中的光照、温度、水分和气压,海洋中的盐分,空气中的氧气、二氧化碳等,如果偏离了动物的生存所要求的水平,就会引起动物的不适,甚至会导致动物死亡。例如,冬天,气温急剧下降,大量的昆虫就会被冻死;如果天旱缺水,许多动物就会被渴死。有些动物随着冬天的到来,气温的下降,还会进入冬眠,停止捕食。所以,在这些恶劣的条件下,动物自身的生存很难保证,捕获食物和繁殖后代的能力毫无疑问会被削弱。
还有动物的竞争(天敌的捕食和对共同食物的争夺)和各种自然灾害等等也都会阻挠动物的捕食。环境中的山脉、森林、河流和其它障碍物也会对动物的捕食起阻碍作用。这些障碍物一方面遮掩了食物,阻扰捕食者获得有关食物的信息,另一方面还会妨碍捕食者捕获食物。
如果我们把环境中不利于动物捕食和繁殖的因素统称为环境阻力,那么由上述的讨论可知,环境阻力越大,动物的捕食力就会越小,反之,环境阻力越小,动物的捕食力就越大。
动物捕食也少不了信息。有经验的老虎及其他动物在捕食猎物时,常常跑到被捕食者的下风去,以便不被猎物发现和根据气味、响声来准确判断猎物的位置而进行捕猎。但是在大自然中,捕食者从来都不可能获得完全的信息。这是因为:
第一,有关食物的信息常常被各种因素(如森林、山脉和其它障碍物)所阻隔。所以,捕食动物来说,要发现食物,必须经常地迁移或游猎,以便不断地发现食物。
第二,被捕食动物常常有意躲避捕食者,使捕食者难以发现它们。一些动物为了逃避捕食者的捕食,常常保持高度的警戒,一有风吹草动,它们就逃得无影无踪。另一些动物在长期的进化中形成了能迷惑捕食者的保护色,使捕食者很难分辨真伪信息。
第三,捕食者发现被捕食者的能力也是有限的。捕食者通过眼、耳和鼻等感官器官获得被捕食者的有关信息,而眼、耳和鼻等器官所能感觉到的范围是十分有限的。由于不能及时准确地掌握有关食物(猎物)的信息,所以动物的捕食能力便大大地受到削弱。
虽然捕食者不可能捕食它们所发现的每一个动物,但研究表明,动物捕获到食物的数量与它们发现的食物数量具有正比关系。
因此,如果用ε来表示环境阻力和信息阻隔程度,并约定ε越大,环境阻力和信息阻隔程度就越高;ε越小,环境阻力和信息阻隔程度就越小,那么就应该有:
F∝1/ε,即动物的捕食力与“环境阻力和信息阻隔程度”成反比。
至此,我们一共列出了影响动物捕食力的四大因素。它们是:(1)物的特性、体质和捕食欲望k;(2)动物的体重Q;(3)环境中食物的分布状况∑(Qi/4πri2);(4)环境阻力和信息阻隔程度ε。
这四大因素并不是各自独立、互不相干的,而是相互联系、相互作用、相互制约的,缺少其中任何一个因素都不行。倘若k=0,表明动物的特性、体质和捕食欲望全都没有,这样的动物根本不可能存在,或者说这样的动物实际上已经死亡了,毫无疑问,此时F=0;若Q=0,即动物没有体重,此种情况表明没有动物或动物不存在,所以也无所谓捕食力,即F=0;若∑(Qi/4πri2)=0,说明环境中无任何食物,显然,此时此刻动物即使有再大的力气它也无用武之地,其F=0;若ε→∞或1/ε=0,亦即动物完全不知道任何有关食物的信息,既看不到食物,也听不到、嗅不到食物,或者环境阻力无穷大,例如由于障碍物(山脉、河流)的阻隔,使得动物寸步难行,不可能得到食物,或者温度特别低,低得动物不能生存(进入冬眠),那么也会有F=0。由此可知,在四个变量之中,只要其中任何一个为零,都有F=0,所以,这四个量对捕食力的贡献不是加和的关系,而应该是乘积的关系。即:
F∝kQ∑(Qi/εi4πri2)(这里εi表示捕食食物Qi的环境阻力和信息阻隔程度)
或F=λkQ∑(Qi/εi4πri2),其中λ是比例常数。
当然,我们可以通过单位的变换和取舍来消除上式中的比例常数,从而得到:
F=kQ∑(Qi/εi4πri2)………………………④
由此可以看出,④式与改造后的万有引力定律②式和库仑定律③式具有完全相同的形式。
这不是偶然的巧合,而是生物与非生物有着某种必然联系的反映。④式的生物学意义是:在某一生态系统中,动物的捕食力与该动物的体重和环境中食物的多少成正比,与环境阻力和食物信息被阻隔的程度成反比,同时还与动物的特性、体质和捕食欲望有密切关系。这便是动物的捕食定律,它是动物界的引力定律,反映了动物的捕食规律,是万有引力定律在动物界的表现形式。
这里需要说明的是,⑤式不仅适用于动物个体,也适用于一个动物群落或动物社群,所以⑤式具有普遍的意义。它从定性和定量两个方面描述了动物的竞争行为和结果。
3.2、吸养力
F=kQ∑(Qi/εi4πri2)公式虽然由动物推导而来,但是,它也适合于植物。只不过在用于植物时,F表示的是植物获得阳光和吸取养分或肥料以及水分的能力,简称吸养力。
通过研究也可以发现,植物的吸养力与植物的重量(Q)和环境中养分(肥料、水分等)或阳光的分布状态(Qi/εi4πri2)成正比,与环境对植物获取阳光和养分(肥料、水分等)的阻碍程度(ε)成反比,与植物的内在特质(k)有关。这就是植物界的引力定律。用公式表示就是:
F= kQ∑(Qi/εi4πri2)