
用译码模型解释色觉机制进化和色盲
根据译码模型,
我们能通过三敏感曲线的变化简单地说明色觉进化(见图15)。
图
15 色觉机制进化图解(横坐标:波长;总坐标:信号强度)请想象曲线R和G逐渐相互靠近以至合二为一, 取名为Y, 我们将看到代表红、绿、青、绛的区域逐渐消失。 进一步, 让B和Y逐渐相互靠拢以至合二为一, 取名为W, 我们将看到表示蓝和黄的区域也逐渐消失, 最终只有白色和黑色区域存在。 我们可以想象颜色视觉是以相反的方式进化的。具有一种锥细胞的人只能察觉两种完全不同颜色:黑和白; 随着W分裂成B和Y, 人能渐渐能分辨出四种完全不同 颜色: 蓝、黄、白、黑。 随着Y分裂成G和R, 人能渐渐地能见到更多完全不同颜色
我们得到结论: n种色敏感细胞可以引起2n种完全不同色觉(对于n=1,2,3)。 当n=4时, 结论看来也是正确的。笔者建立了一个对称的可用于机器人的四原色色觉机制数学模型,其中除了白色” (1,1,1,1)和黑色(0,0,0,0),有14种单 纯彩色”, 它们可以对称地放在一个球面上【9】。 由此我们可以得到一个颜色球它有许多牛顿色盘所具有的性质。
现在我们可以通过视细胞敏感曲线的相互靠近从而更方便地解释色盲。 比如, 全色盲可以说是因为色敏感曲线B,G,R还没有从一种分裂出来; 红绿色盲可以说是因为曲线G和R还没有从Y分裂出来。 从这一观点看, 有些色盲被称之为“红色 盲”或“绿色盲”只是由于Y波峰的波长或长或短。
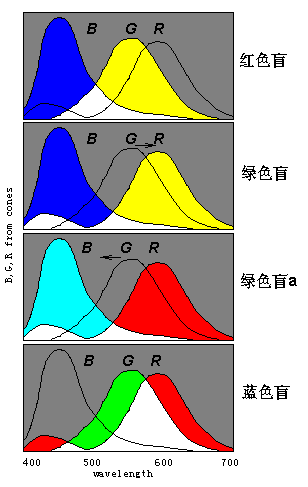
图
16 色盲图解(横坐标波长;纵坐标:信号强度)蓝色盲可以想象B曲线向G曲线靠拢,以至合二为一。
各种色盲也可以通过让模拟电路中两个或三个3DU接收相同的输入信号来模拟。
另外,我们也可以想象绿色盲是因为G曲线合并到B曲线而不是R曲线中了,这样一来,人的色觉就有所不同,不是产生正常人的蓝黄两种感觉,而时正常人的红青两种感觉, 如图中绿色盲a所示。 然而,后面我们将证明,绿色盲a和绿色盲完全等价,不可区分;或者说,一个先天的绿色盲患者不可能知道自己是两种(甚至更多种)绿色盲中的哪一种,其他任何人也不会知道。
上面我们是用物理颜色来表示色觉的,而严格说来,色觉本身是不可表示的,我们只是表示了不同色觉的差异,并没有表示出色觉本身。所以图16中的绿色盲和绿色盲a两种表示完全等价。 和流行的阶段模型不同的是,译码模型是对称的,有三对互补色、六种单纯彩色(unique colors)或感觉信号。我以为理想的色觉机制应该有6种单纯彩色(加上黑白一共是8种――2的3次方)。有人或许认为心理学实验已经证明四种单纯彩色存在, 因而产生六种纯彩色的模型是不合理的。 然而,笔者坚持自己的观点有不少理由:1)说只有四种单纯彩色存在,这并不是最终结论, 有些人仍然坚持说有多于四种单纯彩色存在【27】;
2)生理学实验表明有多于两种彩色互补信号存在于猴类视网膜中【28】;
3)具有对称性的模型更加简洁自然,而对称的色觉模型要求有六种单纯彩色存在;
4)颜色视觉正在进化,我们能想象单纯彩色的数目正在由四种变为六种, 如果说在红色和绛色,绿色和青色之间存在类似性, 这只是由于视网膜中有些构造正处于较低的水平上。 笔者在《广义信息论》中证明,感觉的信息随分辨率提高而增加,人眼看出的单纯彩色越多,色觉能提供的信息就越多。六种彩色是传递更多信息的需要,也是进化的方向。
为什么颜色信号在视细胞阶段以三基色形 式存在而在视神经阶段以互补色形式存在? 我们的回答是:这是由 于经济的缘故。 从物理模型我们能发现, 以这种方式,六种单纯彩色信号能被最为经济地产生和传递。
和流行的阶段模型相比较, 译码模型从信息论的角度和经济性的角度看有着显然的优点;此外译码模型还具有下面优点:
1) 译码模型更加简洁, 对称且不带修正参数;
2) 译码模型中, 从理论上讲, 一个单纯彩色(unique color)可以唯一地(uniquely)产生。 比如, 当(B,G,R)=(0,0,1)时, 只有红度不是0, 白度和其他六个量皆是0。 而在流行的阶段模型中, 除了(B,G,R)=(0,0,0), 白度在任何时侯都不为0。 这就缩小了可能的色觉空间, 这是不好的;
3) 译码模型更和显示在电视屏幕上的颜色混合规律相容;
4) 译码模型可以更加方便解释色盲和色觉机制进化,以及负后象现象(不赘)。